
Evolution

Introduction
Plesiosaurs belong to a larger group of reptiles known collectively as the Sauropterygia. Sauropterygians had a worldwide distribution (Rieppel 1997) and range in age from the Triassic Period to the end of the Cretaceous Period (Benton 1990a). The fossil record of sauropyerygians is relatively rich because aquatic environments have high rates of deposition and so organisms that inhabit water have a relatively high preservation potential. Consequently, “sauropterygians provide the most complete evidence of the sequence of events that leads to a specialised aquatic way of life” (Carroll 1988). However, the absence of articulated remains in the Late Triassic (Carroll 1988, Storrs 1993), has limited our knowledge of the “nothosaur”-plesiosaur transition.
There have been numerous convergence and extinction events in sauropterygian history (Bakker 1993). Detailed analysis of the cranium of plesiosaurs has demonstrated that short-necked and long-necked plesiosaurs evolved independently several times. For example, the short-necked polycotylid plesiosaurs, including Dolichorhynchops osborni, were once regarded as pliosaurs, but are now known to be a separate lineage (Carpenter 1997). As such, the evolutionary history of plesiosaurs is more complicated than previously anticipated, and this is an active area of research today.
Ancestors
The exact evolutionary origins of sauropterygians is unresolved (Taylor 1989). Primitive diapsid eosuchian families, such as the younginoid family Tangasauridae, are the most likely close ancestors (Carroll 1988). Members of this group show modifications of the tail (lateral flattening) for an aquatic mode of life, which also puts them in an appropriate ecological setting.
Claudiosaurus from the Upper Permian of Madagascar was once considered a sister taxon to the Sauropterygia (Carroll 1988, Storrs 1993, and Piveteau 1955 in Storrs 1991). Similarities between this genus and sauropterygians include an elongate neck, paddle-like distal limbs, and a euryapsid condition in the skull (lack of a lower temporal bar).
The palate of Claudiosaurus also resembles that of sauropterygians, especially the closed condition of the palate present in pachypleurosaurs and “nothosaurs”. They both lack a transverse flange of the pterygoid, and have reduced suborbital fenestrae and interpterygoid vacuities. The upper temporal opening is smaller than the orbit, as in pachypleurosaurs (Carroll 1988). The sternum shows little ossification (Carroll 1988), a common feature in aquatic reptiles known as “aquatic neotony” (Ricqles 1975, Rieppel 1987b in Storrs 1993). However, these characters may have evolved independently.
The closest known relatives of plesiosaurs are the pistosaurids (and other non-plesiosaurian pistosaurians/pistosauroids). This group of Triassic sauropterygians is known from several genera around the world: Pistosaurus from Germany, Augustasaurus from the USA, Bobosaurus from Italy, and Yunguisaurus from China. The pistosaurids can be regarded as intermediate between true plesiosaurs and ‘nothosaur-grade sauropterygians. They are plesiosaur-like in many aspects but lack some of the extreme adaptations to marine life seen in plesiosaurs.
Size
Change in body size and proportions in sauropterygians occured as a result of speciation as they adapted into a variety of niches. In general, the size of individual animals becomes greater over geological time, and derived clades tend to be large than primitive ones. The tiny pachypleurosaurs range from 20 to 100 cm long, “nothosaurs” range from approximately one to four metres long, and the more derived plesiosaurs (pliosaurs and elasmosaurids) reached lengths of 15 metres or more.
Cranial data
There is widespread agreement that the absence of the lower temporal opening is in fact an unusual variation of the diapsid condition rather than being a unique “euryapsid” condition. (Taylor 1989). However, the position of sauropterygians within the Diapsida is less well constrained. Rieppel (1989) (in Taylor 1989) considered them to be either primitive diapsids yet to evolve the second opening, or to have lost the lower temporal opening via secondary derivation.
The upper temporal opening of the pachypleurosaur head is smaller than the orbit (Carroll 1988) a plesiomorphic state, not a defining factor of the group (Storrs 1993) and the upper temporal bar is strong. The temporal opening is larger in the nothosaurs and in plesiosaurs increasingly so (Carroll 1988). The palate of pachypleurosauria and “nothosaurs” has posterior and medial extentions of the pterygoids, the interpterygoid vacuities are completely closed and ventrally the base of the braincase is covered. This is considered an extension of the condition seen in Claudiosaurus (see below) (Carroll 1988). The pachypleurosaur skull is characterised by loss of the ectopterygoid (Rieppel 1989 in Storrs 1993) in addition to the reduction of the supratemporal and an impedence matching middle ear.
A distinguishing character of the plesiosaur skull is the loss of the nasal bones, a synapomorphy of Pistosaurus (Carpenter 1997). This is however slightly unreliable as misidentification of the prefrontals is common (Carpenter 1997) and O’Keefe (2001a) proposed that this is only synapomorphic for the Plesiosauroidea (i.e. Pliosauroids posses nasals). The palate of Pistosaurus is basal (Sues 1987) indicating that the plesiosaur lineage perhaps broke off before the pachypleurosaurs (Carroll 1988). Plesiosauroids tend to have upwards facing eyes while pliosauroids are laterally placed (Massare 1988). This may have implications for feeding strategies.
In pliosauroids the mandibular symphysis may be short with 5-6 caniniform teeth or long with 10-12 caniniform teeth (Tarlo 1960). Primitive elasmosaurs are identified by the presence of contacting frontals (Wegner 1914 in Carpenter 1997). In the Cretaceous elasmosaurids and polycolylids, the pinneal foramen is dorsally closed, a synapomorphy, and in latest Cretaceous genera the frontals are separated by the premaxilla and form the dorsal rim of the orbit.
In plesiosaurs, the lachrymal is only present in pliosauroids and is absent in elasmosaurids and polycotylids (allegedly absent in plesiosaurids) (Carpenter 1997), however, this may actually be the nasal bones (see above). In contrast to other Sauropterygia, the elasmosaurid jugal forms most of the edge of the orbit. In the elasmosaur Libonectes, a small fenestra indicates a remnant of the suborbital fenestra (Carpenter 1997). The exoccipital-opithotic bone is not part of the occipital condyle in all plesiosaurs except Cryptoclidus and pliosaurs indicating an early divergence of this group. Synapomorphies between the elasmosaurids and polycotylids include the presence of a vomeronasal fenestra, expansion of the pterygoids into plates below the braincase and loss of the pineal foramen and stapes.
In pliosauroids of Upper Jurassic sediments of Britain, the younger Oxfordian genera have circular cross-section teeth (patterns may be used for distinguishing species) while in the Kimmeridgian, all pliosaur teeth are trihedral in cross section (Tarlo 1960).
Postcranial data
Pectoral Girdle
The pachypleurosaur and “nothosaur” pectoral girdle is poorly ossified. The reduced ventral portion of the pachypleurosaur girdle reflects their basal position (Carroll 1988): neither the clavicles or the scapulae are expanded ventrally as they are in the “nothosaurs” and to an even greater extent in the Plesiosauria. However, derived features of the pachypleurosaur pectoral girdle include the massive scapula and the lack of posteromedian processes on the interclavicle. The coracoids are larger in “nothosaurs”, a reflection of their increased reliance upon limbs for movement (Storrs 1993).
There is a similarity between the pectoral girdles of adult plesiomorphic taxa such as Pachypleurosaurs and “nothosaurs” and the juvenile form seen in plesiosaurs (Storrs 1993, Carroll 1988). This is evidenced by the similarity between the pectoral girdles of juvenile Cryptoclidids, for example Cryptoclidus, with the adult forms of related but more primitive sauropterygians e.g Neusticosaurus and Ceresiosaurus. The limb girdles of plesiosaurs are ventrally expanded but the vertical components are reduced (Storrs 1993). This is evidence of possible heterochrony within the sauropterygian lineage. Such peramorphosis (addition to form) also occurs in plesiosaur limbs. Typicaly elasmosaurids have a midline scapular bar. Further more, the pectoral girdles of placodonts such as Placodus show even less development, further evidence for their perceived primitive position.
Pelvic Girdle
This is reduced in the basal sauropterygians as they relied on asymmetrical undulatory motion, but expanded again in the plesiosaurs whose mode of locomotion involves the symmetrical use of limbs (Carroll 1988).
Vertebrae
Pachypleurosaurs and “nothosaurs” have lost the intracentra except at the base of the tail and to the anterior of the atlas-axis. The number of sacral vertebrae in Sauropterygia ancestors is two, in pachypleurosaurs three, in “nothosaurs” six (Carroll 1989). The neural spine of pachypleurosaurs is low and the neural arches are broad and flat and the tail is long, often longer than the rest of the body (Sues 1987). This contrasts to the tall broad neural spines and sub-rectangular narrow neural arches, flat vertebrae faces and short laterally compressed caudals observed in “nothosaurs” (Sues 1987). The thoraces of “nothosaurs” and plesiosaurs are rigid (Carroll 1989) but those of pachypleurosaurs are less so. Densely packed gastralia occur in the “nothosaurs” (Storrs 1993). This is also a feature of the Plesiosauria, a derived character as indicated by the lack of accessory vertebral articulations (Storrs, 1993). In pliosauroids the cervical vertebrae are distinguishable from other sauroptertgians as well as between inclusive species but unusually not between genera (Tarlo 1960). It is the presence or absence of a ventral keel on the anterior cervical vertebrae which is an important distinguishing feature of pliosauroids (Tarlo 1960). Pachyostosis is common in the ribs and vertebrae of many pachypleurosaurs e.g Neusticosaurus as a mechanism for achieving neutral buoyancy. This is rare in plesiosaurs but such thickening of bone has been observed in the pliosauroid genus Pachycostasaurus dawni from the Middle Jurassic (Cruickshank et al. 1996).
Limbs
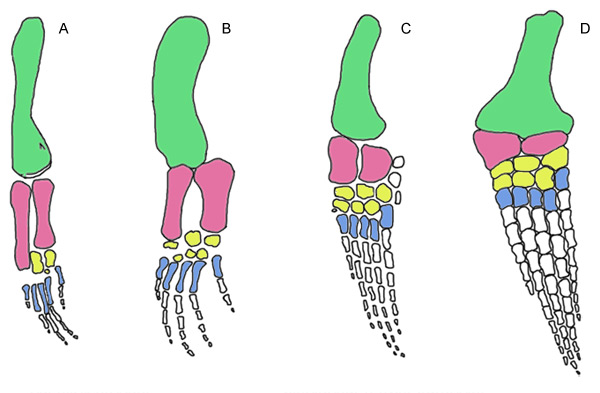
Evolution of the forelimb in sauropterygians, oldest to the left. A. Pachypleurosaur, B. ‘Nothosaur’, C. Early Jurassic plesiosaur, D. Late Jurassic plesiosaur. Note the increased number of phalanges, the closer spacing between the bones, the distal broadening of the humerus (green), the shortening of the epipodials (pink), and the increasingly wing-like overall shape in more derived forms. Propodials shaded green, epipodials shaded pink, mesopodials shaded yellow, metapodials shaded blue, phalanges (finger bones) and accessory ossicles shaded white. (Redrawn and modified from Storrs 1993)
The limbs of sauropterygians become increasingly modified for a fully aquatic life style. In pachypleurosaurs the humerus is large and the distal elements are reduced. The condition in “nothosaurs” is similar but some forms e.g. Ceresiosaurus show some hyperphalangy, increase in the number of finger bones in each digit. (Carroll 1988, Storrs 1993). The number of actual digits is always five, but the trend towards increased number of phalanges continues in plesiosaurs. The functional surface of the ‘nothosaur’ forelimb is increased by distal flattening and broadening of humerus, and this trend increases in plesiosaurs, which have greatly expanded propodials (humerus and femur). The epipodials become increasingly short; this reduces flexibility of the limb as a whole, and makes it a more efficient flipper. In the plesiosaurs, the limbs are modified into wing-like hydrofoils, which taper distally. (Storrs 1993, Sues 1987). Their flexibility was limited furher by the close spacing of the elements and by the alternating sequence of the phalanges. The only functional joint in plesiosaurs was the shoulder/hip joint.
Plesiosauroids can be distinguished from pliosauroids by the relative size of their fore- and hindlimbs: the hind limbs in plesiosauroids are shorter, whereas they are larger in pliosaurs (Bakker 1993, Storrs 1993). The hind limbs are longer in all sauropterygians with the exception of plesiosauroids (Sues 1987b).
Functional evolution
Sauropterygian locomotion has been studied numerous times (see Robinson 1975, Storrs 1993). It is likely that the phylogeny of the Sauropterygia parallels an increasing adaptation to a fully aquatic existence via increasingly economic swimming regimes (Sues 1987). The underwater flight employed by plesiosaurs is more economic than the undulatory locomotion employed by pachypleurosaurs and as such, the change in body proportions can be attributed to the gradual surpression of lateral truncal movement. This must have involved a radical shift in the central nervous system (Storrs 1993). The reduction of appendicular girdle and limbs in the pachypleurosaurs may be a response to decrease drag (Storrs 1993). A strong ancestral constraint is associated with this transformation in locomotory repertoire, and this is poorly understood.
